Why Lips?
One of the things that seems to get lost in the lips discussion is the evolutionary reason for lips. Unfortunately, little seems to be explicitly published on the evolutionary pressures that would favor lips. However, we can glean a few likely reasons why extant lipped taxa have them, and see if these reasons apply to dinosaurs.
Hypotheses I have seen proposed so far in online discussions are:
- Prevent tooth decay/dessication
- Protect teeth from damage
Neither of these are particularly compelling, since it fails to explain loss of lips in crocodiles (crocodiles spend part of their time out of water and are hyper predators that often risk damage to their teeth). It also fails to explain why sem-aquatic lizards (water monitors, marine iquana, etc.) have not lost their lips.
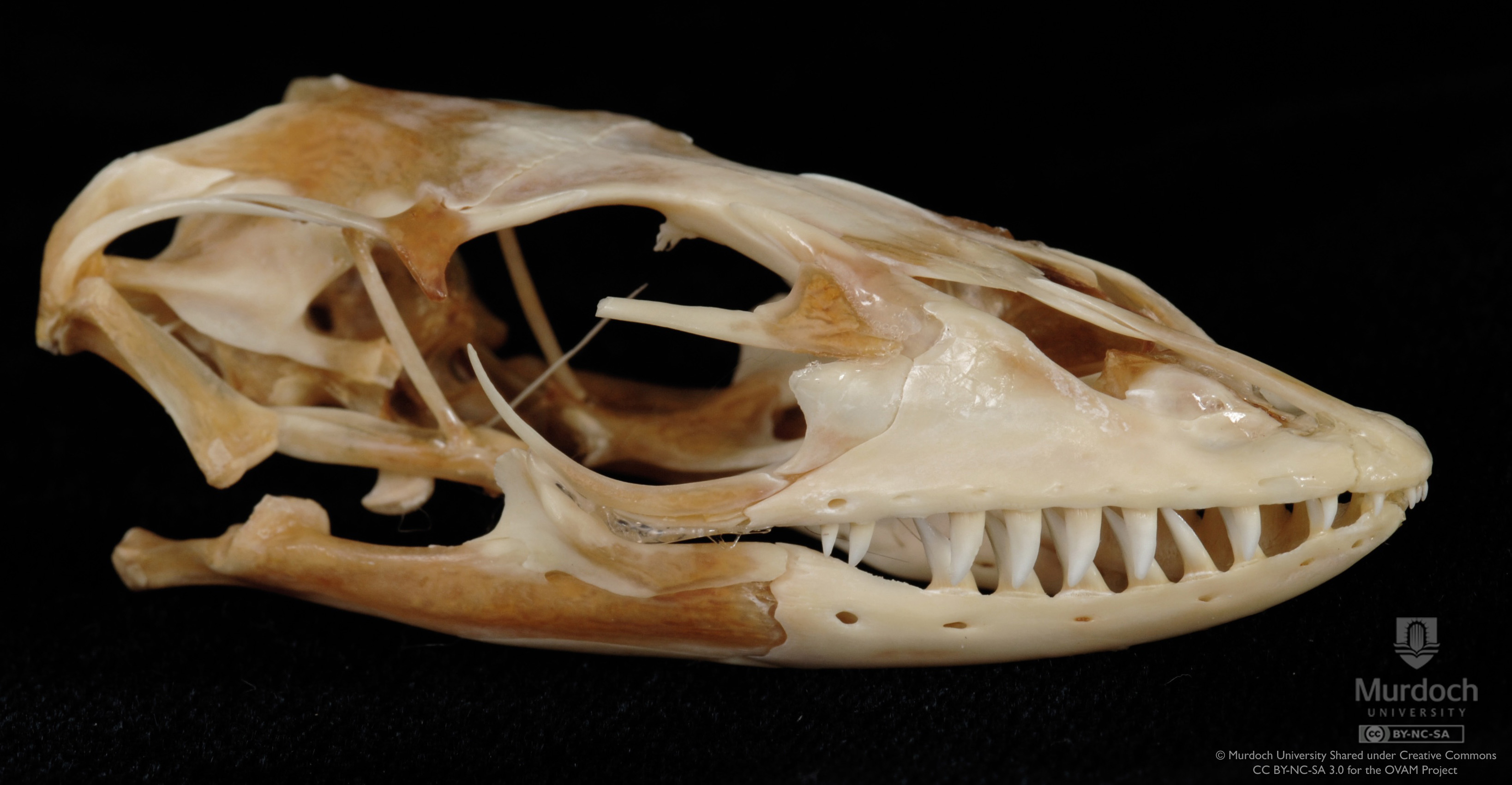
Also, unlike mammals and many lepidosaurs, but similar to crocodiles, dinosaurs repeatedly shed and replaced their teeth. In addition, and unlike crocodiles, most dinosaur species have rows of extra teeth right behind, ready to quickly take the place of any fallen teeth. This would mean there would be even lower evolutionary pressure to evolve or retain lips (if they inherited them from a common ancestor) for purposes of protecting teeth. Furthermore, the gingiva in crocodilians are very tough and thick, similar to what is seen as preserved in a specimen of Camarsaurus and implied by other sauropod isolated tooth rows (Wiersma et al, 2016, 2019; Canudo et al, 2018)..
I think it is more likely that lips in mammals and lepidosaurs evolved due for the following reasons (in order of relative importance):
- in mammals,
- aiding in the suckling of offspring (lips help form a seal to efficiently consume milk)
- acquiring & digesting food
- this would be most important in small insectivores in order to help prevent prey escaping, and in herbivores in manipulating challenging food
- increased surface area for saliva production would help while chewing food for predigestion (and regurgitation in some species)
- social signals
- in lepidosaurs,
- acquiring & digesting food
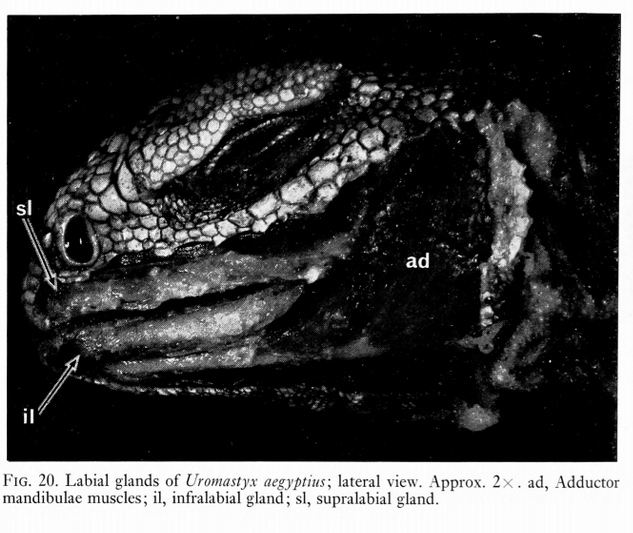
- because many species tend to swallow food whole, lips would aid by increasing the surface area of saliva production, enabling easier swallowing and predigestion
- in venomous species, aid in envenomation
- display
- acquiring & digesting food
Interestingly, crocodiles do not generally eat their prey whole nor do they chew, but rather tear off pieces in chunks and then swallow the smaller pieces. As regards lips being a requirement to safeguarding the gums, crocodiles have extensive, toughened gums (Putterill, Soley, 2003). These are so tough that if the teeth are pulled from the skull, the dental row can stay together.
Birds will swallow prey whole, but also have a gizzard that assists in processing food, as well as tearing off pieces of fleshes and then swallowing the smaller pieces whole.
Based on fossils that show the gut contents, some predatory dinosaurs did swallow food whole and coprolites seem to indicate that they also ripped of chunks of flesh and bone, rather than eating their prey whole. This means that there was some evolutionary pressure to acquire lips in terms of oral processing, similar to large lizards (e.g., Komodo dragons).
Overall, it would seem dinosaurs had less pressure to evolve lips, as they seemed to consume prey quickly with relative large mouths and long digestive track, did not need to nurse or suckle young and had high tooth replacement rates.
Why Beaks?
The other interesting topic is, if dinosaurs did have lips, why would they lose or reduce them repeatedly in favor of beaks?
It is commonly assumed that beaks evolved as a weight-saving feature in volant clades. However, tooth loss or reduction occurs repeatedly in favor of beaks in many non-volant archosaur clades:
- shuvosaurs,
- aetosaurs,
- silesaurs,
- therizinosaurs,
- oviraptorids,
- Limusaurus,
- ornithomimosaurs,
- alvarezsaurs,
- in the lower jaw at least once in Ornithischia, and probably multiple times in the upper jaw (since most of the members of the major clades have basal members with some teeth in the premaxillae))
Beaks occur in pterosaurs as well, but occur in both toothed and toothless species (e.g., Rhamphorhynchus).
In addition, a partial beak may occur in Bonitasaura and a tough keratinous covering may occur in rebbachisaurids (e.g. Lavocatisaurus) and Camarasaurus (Wiersma et al, 2016, 2019; Canudo et al, 2018). Particularly interesting is the case of Camarsaurus where some of the external oral structure was preserved, and the authors indicate that the texture is similar to that seen preserved on the plates in Stegosaurus ( Siber and Urs Möckli, 2009). It appears then that beak-like structures or full on bird-like rhampothecae occur in all the major dinosaur clades.




The beak-like structure seen in some sauropods is said to potentially offer support for the teeth and analyses in beaked theropods show reduction in stress too (see image below).
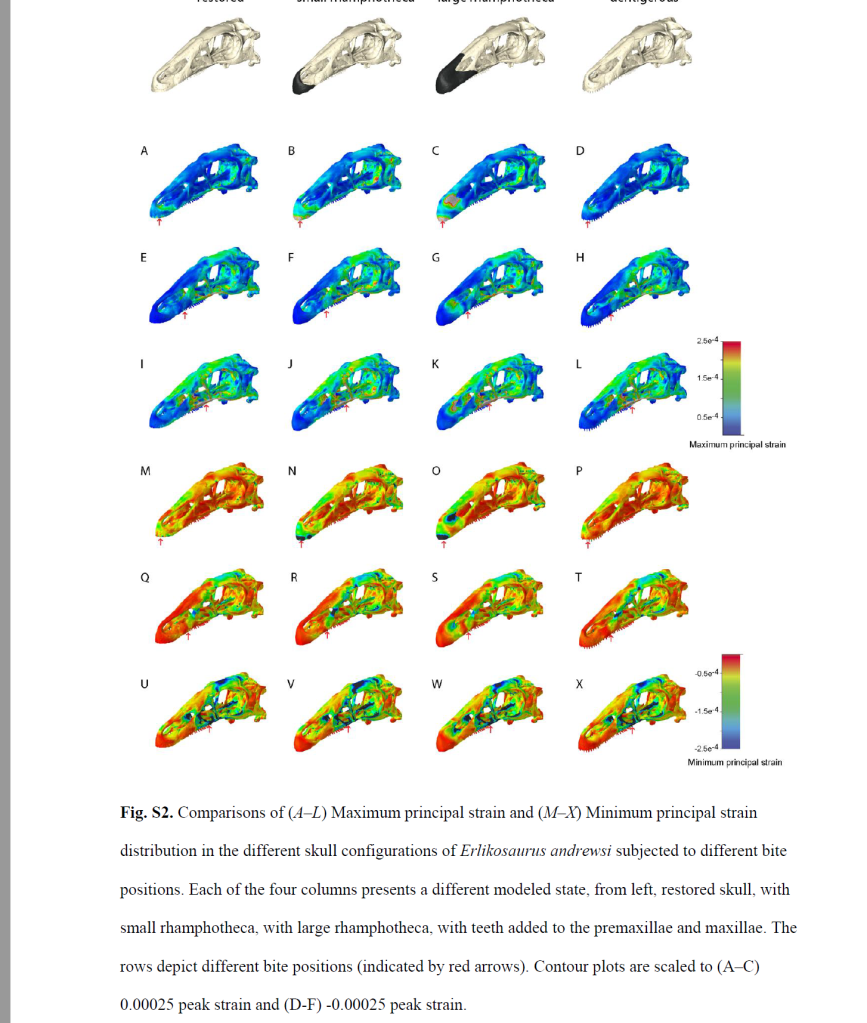
Also interesting is, that none of the volant mammalian taxa (e.g., bats and gliding mammals), show any hint of evolving a beak. Bats are second-most speciose clade of extant mammals, and all have teeth.
This repeated evolution of beaks is strange, considering how rarely it occurs outside of Archosauria. None of the lepidosaurian clades (with the exception of the sphenodonts, if you can call what they have a beak) have evolved keratinous beaks, as well as none of the mammals (exception: if you consider the “bill” in the platypus and echidnas to be a beak, although it is still rubbery and somewhat flexible), and possibly only one lissamphibian group (the sirens).
If evolving from lips to beaks was not a barrier to evolution, one would expect it to occur more often in the highly diverse and speciose mammalian and lepidosaurian clades.
A developmental evolutionary constraint?
In his 1997 paper, “The evolution of beaks in reptiles: a proposed evolutionary constraint”, M.S.Y. Lee discusses developmental hurdles to evolving a beak. He primary focuses on the presence of a caruncle early in ontogeny as a requirement for evolving a keratinzed beak. The caruncle occurs in these clades:
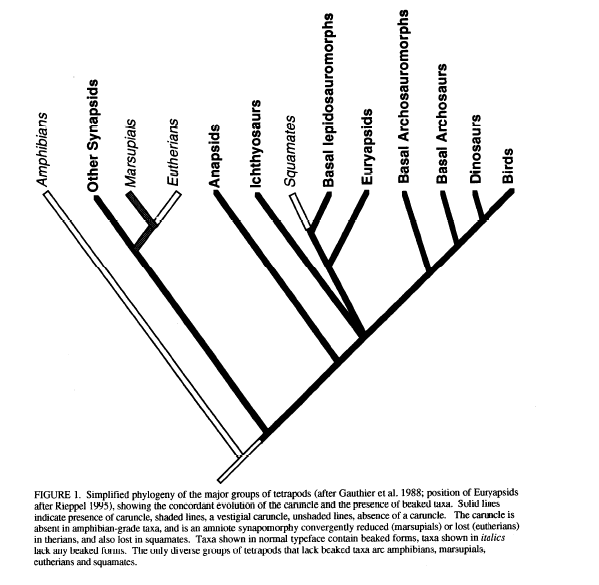
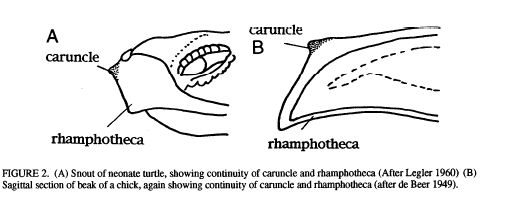
Lee proposes the reason why certain clades have caruncles and others don’t is due to the evolution of viviparity. E.g., the caruncle serves as the egg-‘tooth’ (not actually a tooth) in hard-shelled taxa, whereas an actual tooth serves the same purpose in squamates and egg-laying mammals. This might explain why the earliest mammals, monotremes, started to evolve a beak-like structure, whereas no other mammalian clades did. Furthermore, the requirement of having a caruncle to break out of a hard egg-shell is probably one of the most difficult constraints to overcome evolutionary, since it is extremely closely related to successful reproduction.
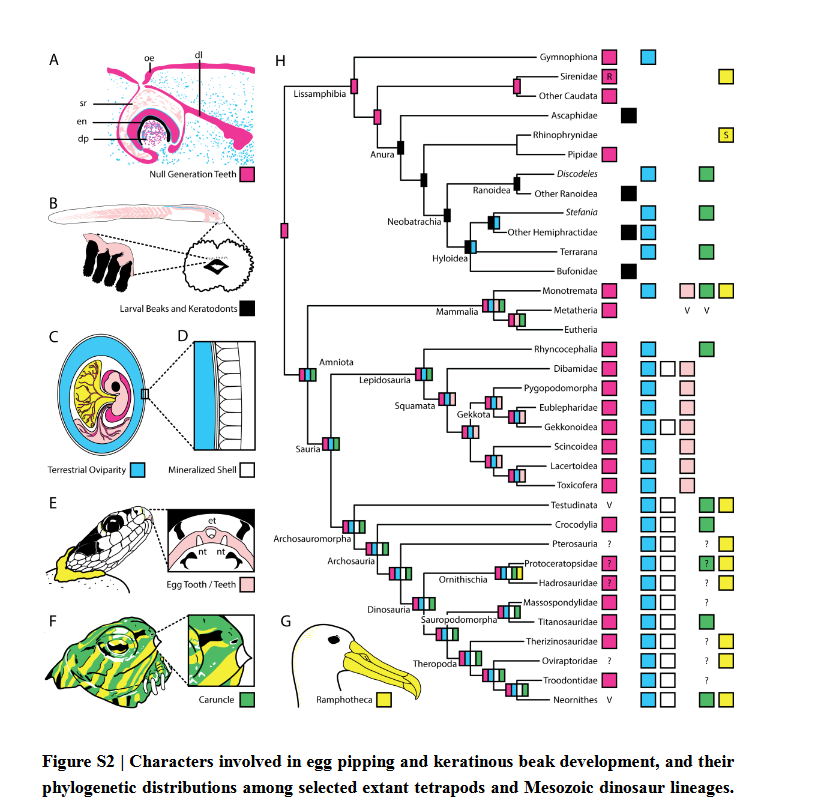
We know that dinosaurs had caruncles for their egg-‘teeth’ instead of actual teeth, based on well-preserved titanosaur embryos which show a horn-like structure.
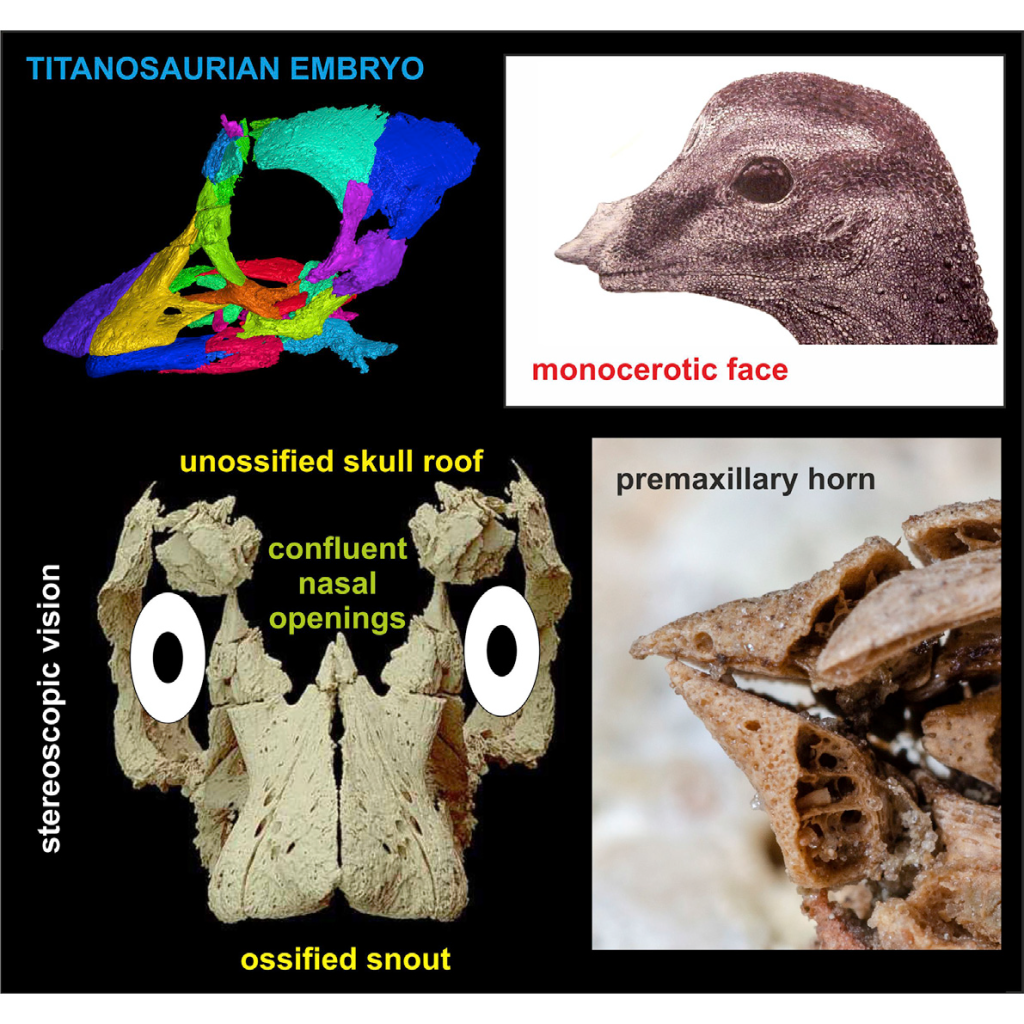
The “egg-tooth” seen in dinosaurs, as has been preserved in sauropods, is similar to that seen in crocs and birds and not the kind of egg-tooth (which is an actual, temporary tooth) seen in mammals and squamates, and some evidence for a horn-like structure (García ,2007 ; Kundra ́t et al 2020).
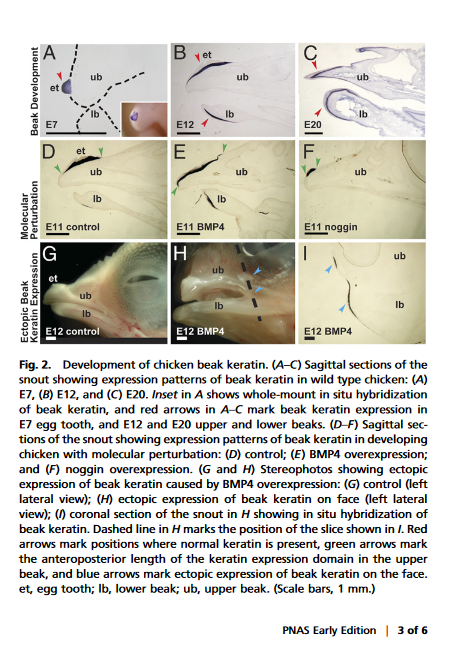
Interestingly, the beak precursors, caruncles, are absent in both therian mammals and squamates (Hieronymus, Tobin. ,2010). This is highly suggestive that there is come developmental antagonism that prevents beaks from evolving in lineages without it. There is no embryological evidence that I am aware of that lips appear in the embryos of turtles, birds or crocodiles and that beaks evolved from them, which would mean your hypothetical scenario of lips being gradually lost in favor of beaks is with out any basis embryologically or genetically as far as I can tell.
Wang, et al (2017) talks about the beak development in adult Limusaurus from toothed juveniles matches the evolutionary developmental scenario of modern bird beaks, which basically shows that the epidermal cells all become cornified during development.

What is also interesting is there is no precedent for having both lips and beaks, unlike beaks and teeth which occur together in pterosaurs and dinosaurs.
So the question is: does the caruncle also serve as an evolutionary restraint for evolving lips? I don’t think we can answer this right now definitely, and the evidence is mostly circumstantial, but I would tentatively propose the answer is ‘yes’ based on the evidence deduced so far. It would appear that keratinized epidermal layers prevent the evolution of lip-like structures by preventing caruncle ‘egg teeth’.
Summary
My reasoning for the absence of lips in dinosaurs is as follows:
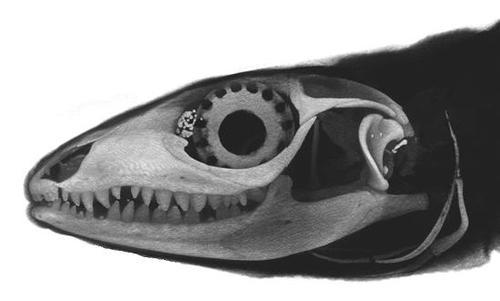
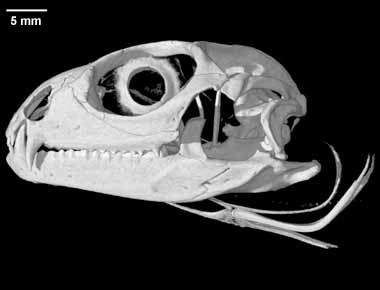
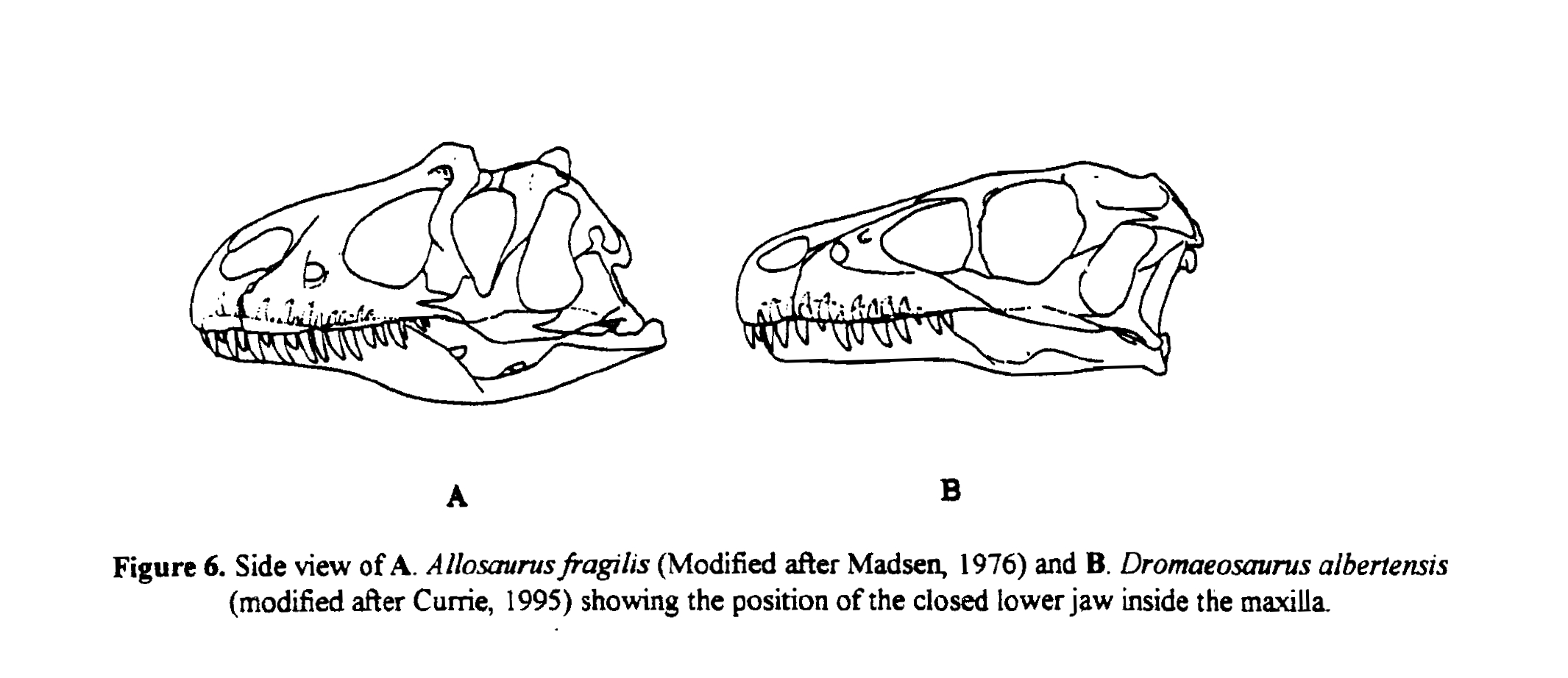
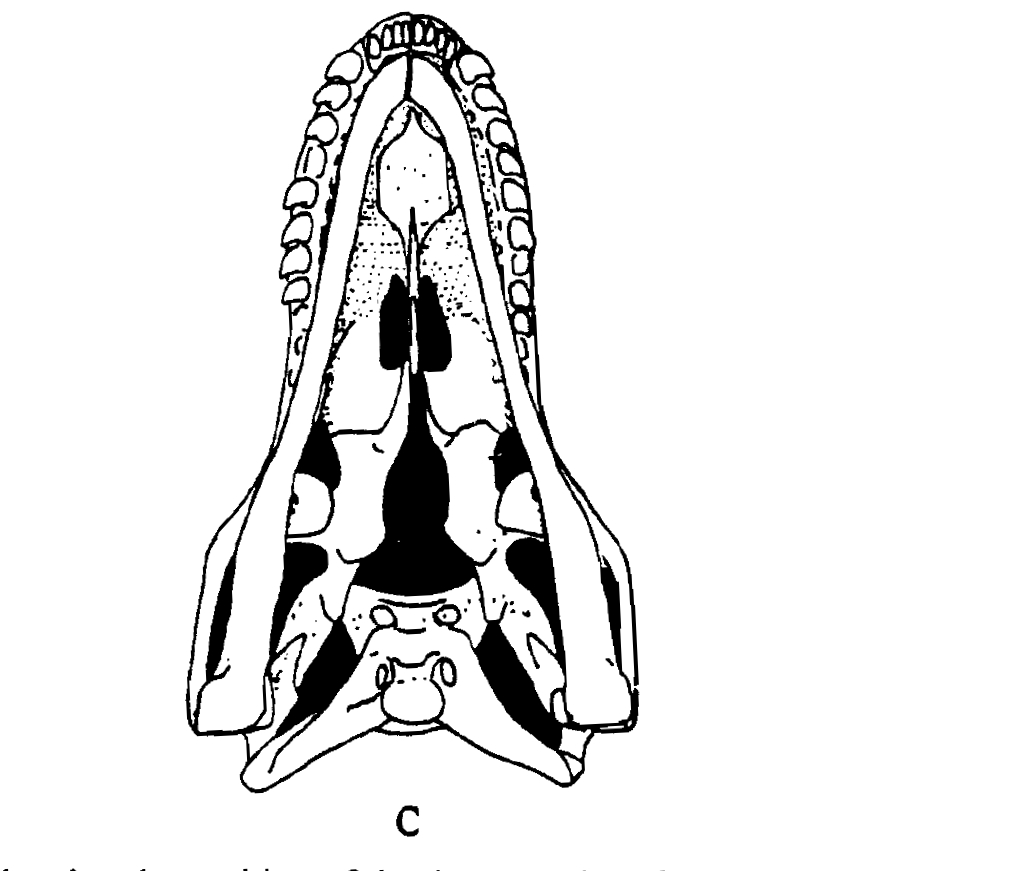
(1) The osteology around the margins of the mouth area in dinosaurs is at least as similar, if not more so, to non-lipped living terrestrial vertebrate taxa than it is in lipped taxa, as well as showing enormous variation within all of those clades that do have lips and beaks (see images below with annotations). This falsifies the hypothesis that there are strong, unamabigous osteological correlates for lips or other extra oral structures.
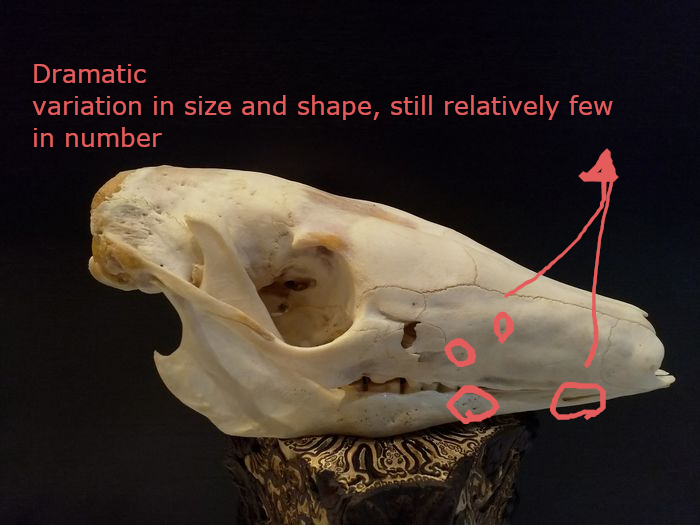
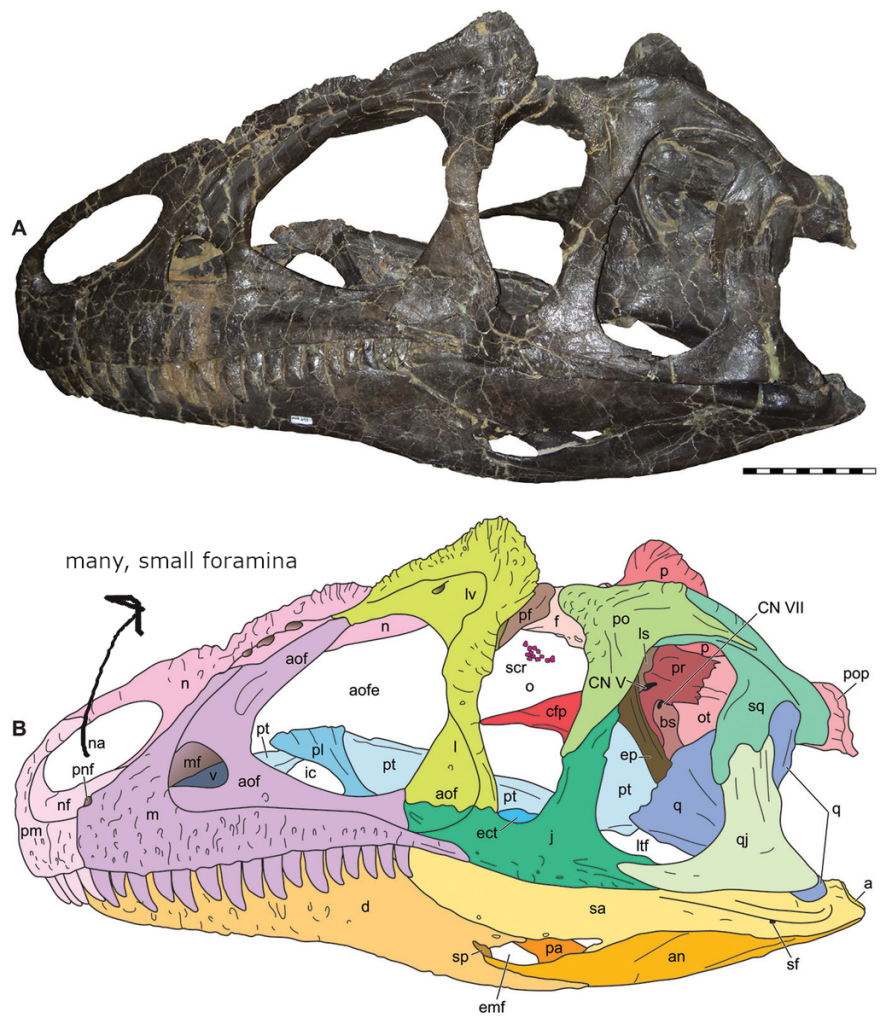
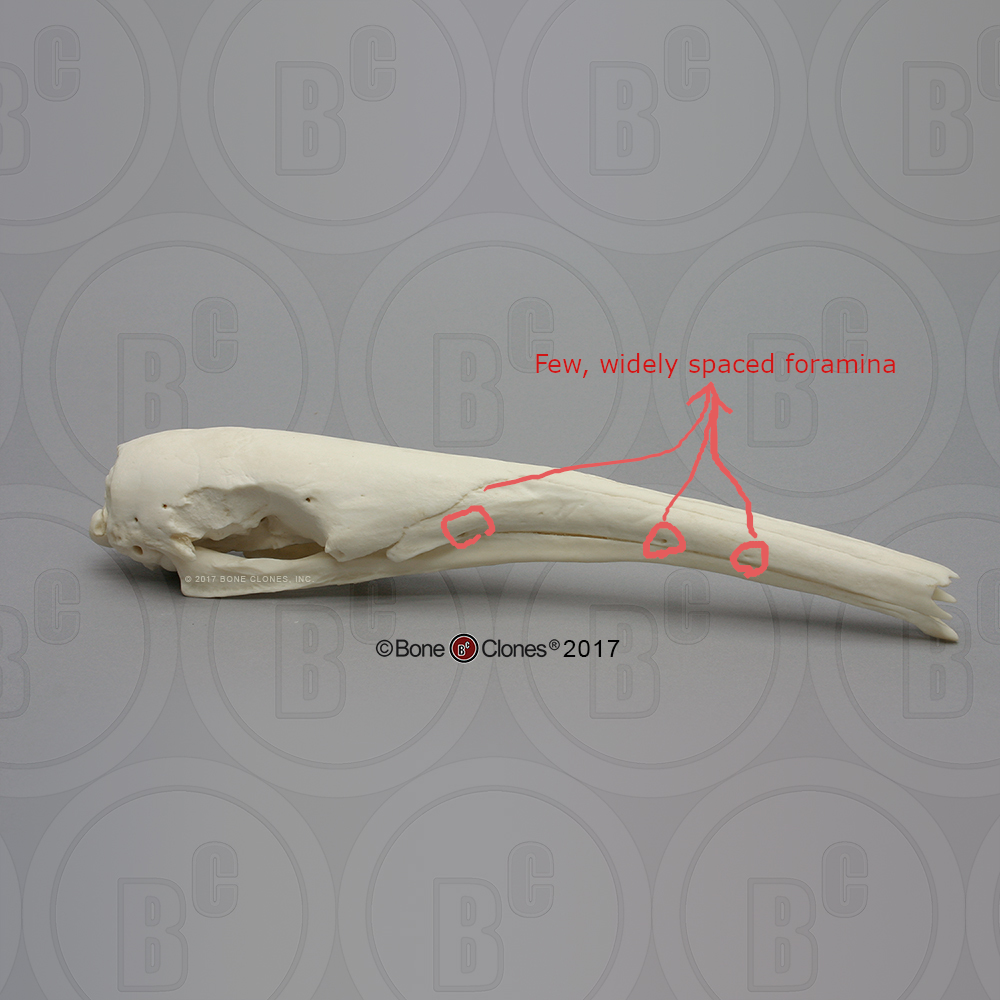
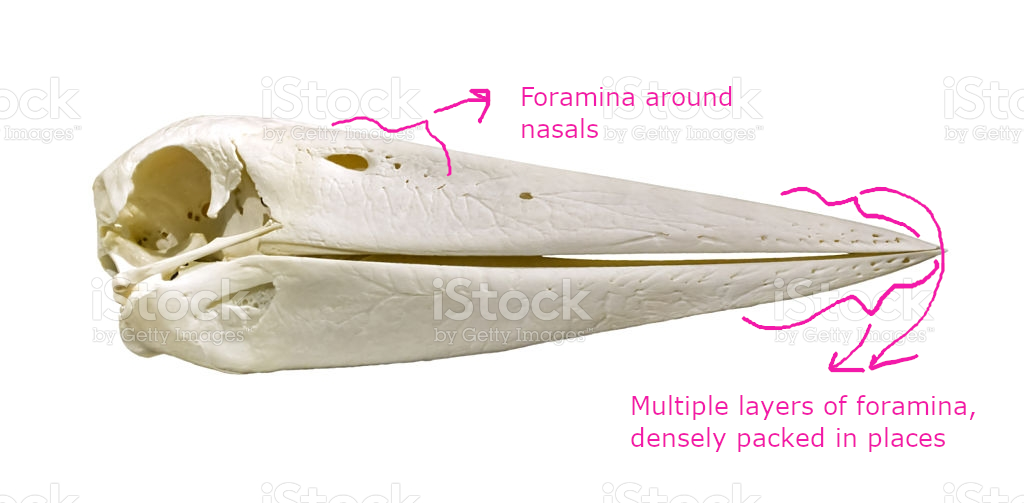
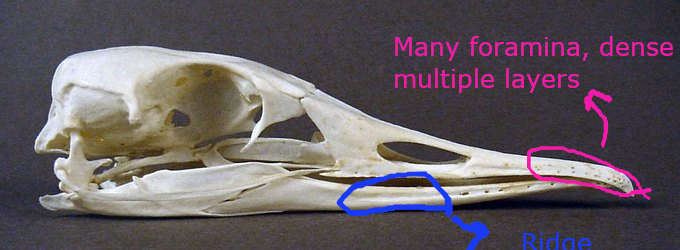
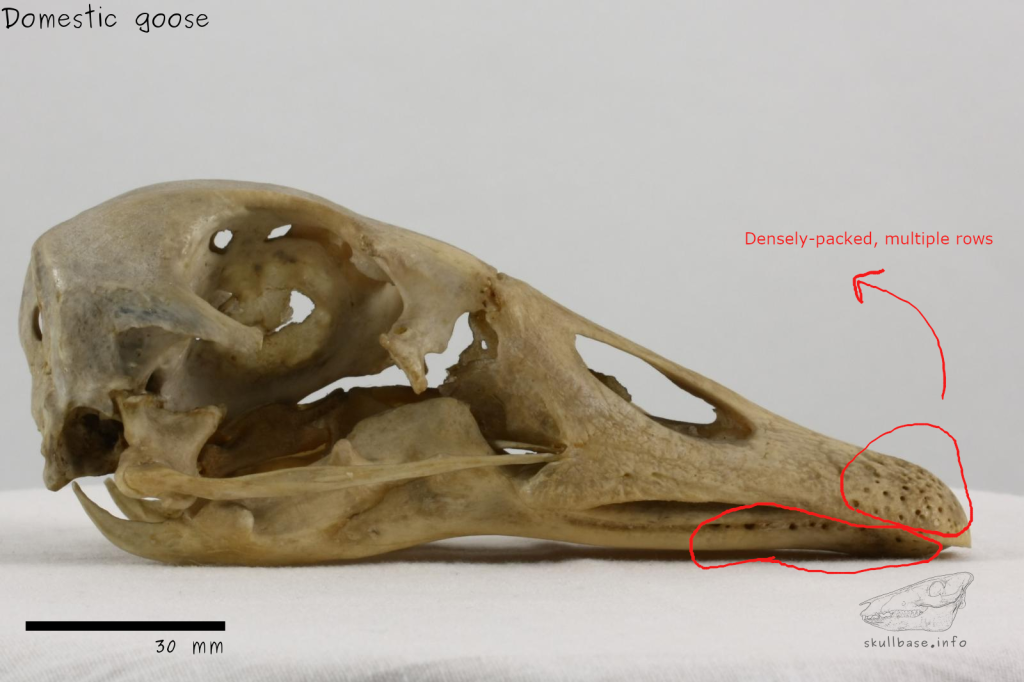

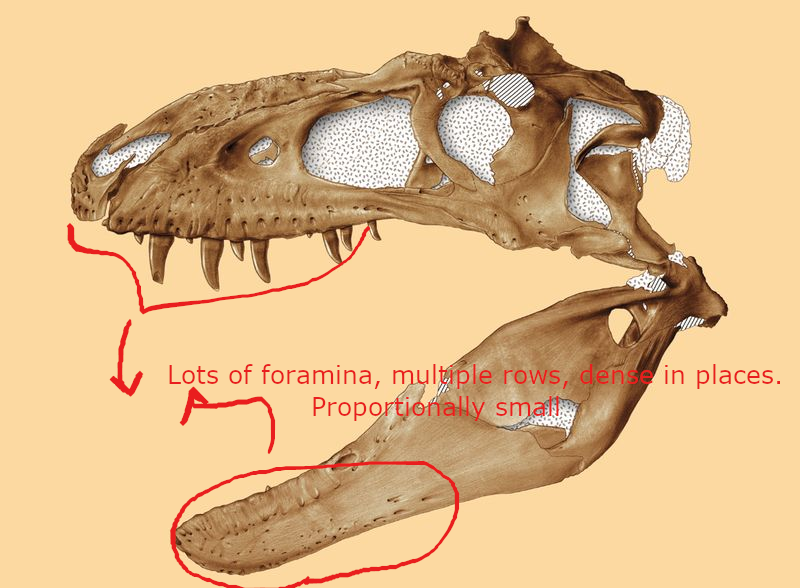
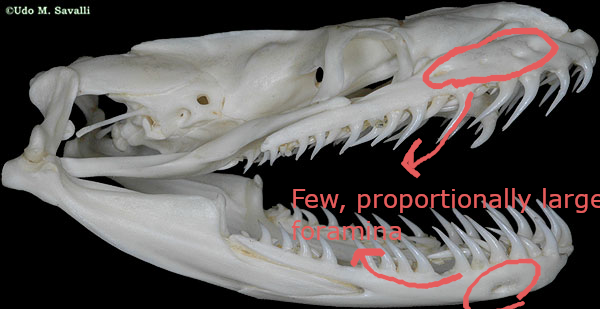
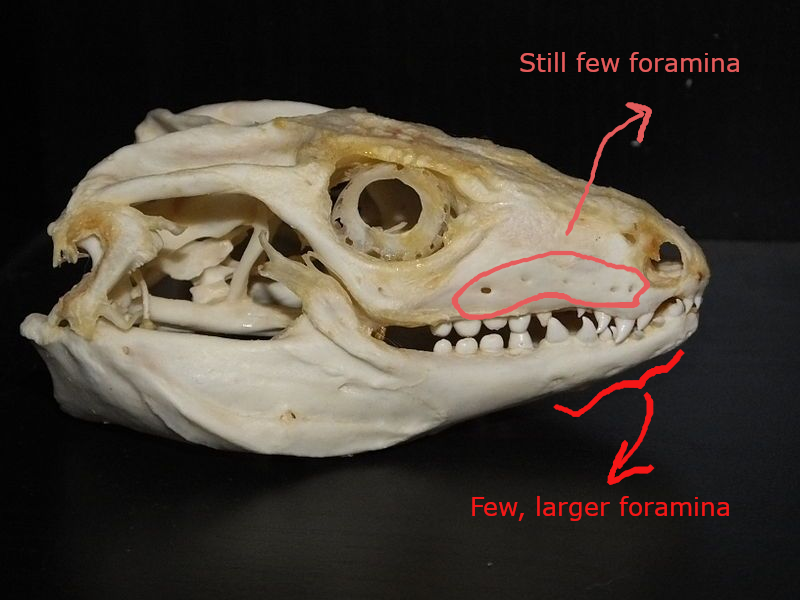
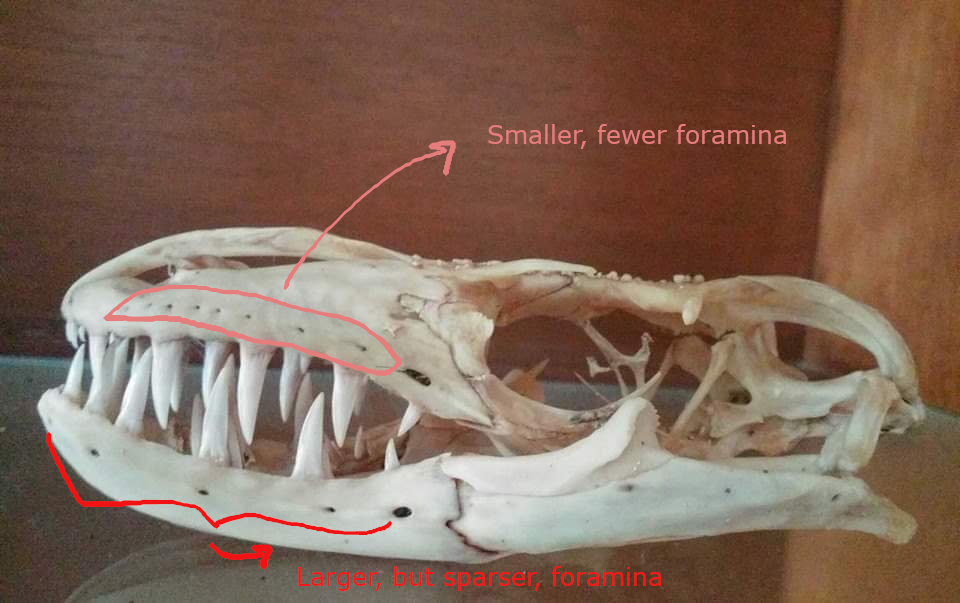
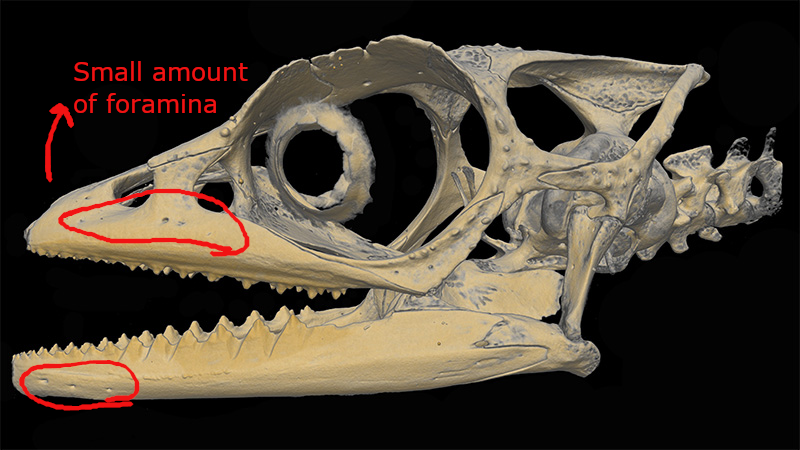
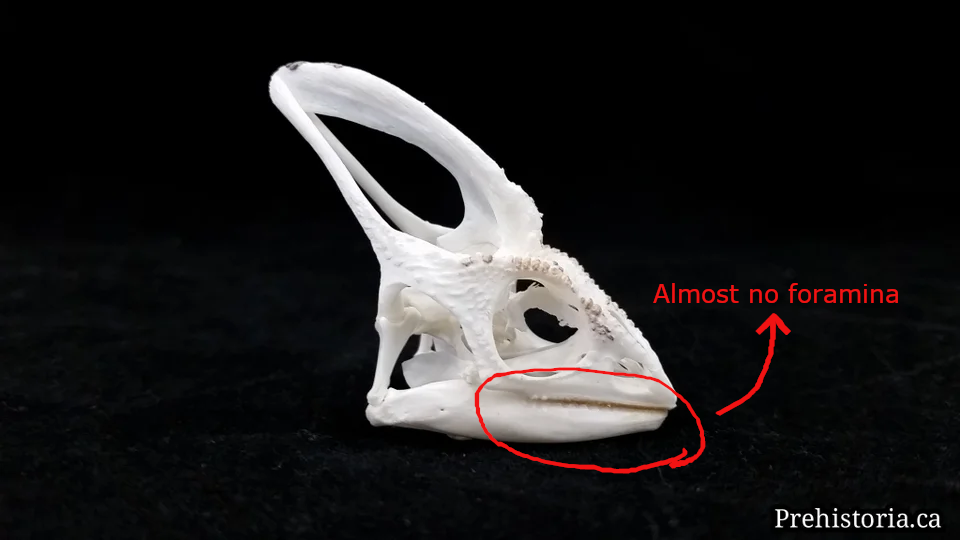
(2) all the evidence we do have for extra oral structures happens to be of hard, keratinized structures on the skulls around the oral margins in dinosaurs, and lack of preserved structures indicating lips. Furthermore, we have evidence of repeated evolution of true keratinous beaks in many other dinosaur subclades, and sister groups (e.g., silesaurs, pterosaurs) and other archosaurus (Effigia, Shuvosaurus). That, combined with the evidence from the EPB, suggest that the most parsimonious reconstruction would be lipless dinosaurs with the basal condition being hard, keratinous oral structures that evolved into true beaks in many dinosaur subclades.
(3) Embryologically, there are more similarities to birds and crocs
(4) Little to no adaptive reason to have had lips, since evidence suggests that dinosaurs replaced their teeth multiple times per year, and since dinosaurs had very different feeding mechanisms from lepidosaurs and mammals there is no reason they would have evolved analogous structures. The evidence so far suggests that they had thick, tough gingiva “gums” like that seen in crocodiles. Furthermore, the lips in lepidosaurs are associated with producing various types of non-venomous saliva to aid in prey capture (Zaher, H. et al, 2014), which evolved into venom in several clades of lepidosaurs. In mammals, lips aid in suckling young – which would be a strong evolutionary pressure.
In summary, the difference in life histories, food consumption habits and evolutionary distance in animals we know have lips, along with the osteological data and the preserved fossil oral strucutures we do have for them, weighs heavily against the idea that lips were common in dinosaurs. Of course, new evidence could come to light and be described that shows otherwise, but until then I think this is the best interpretation given the data at hand.
Refs—
PUTTERILL, J.F. & SOLEY, J.T. 2003. General morphology of the oral cavity of the Nile crocodile, Crocodylus niloticus (Laurenti, 1768). I. Palate and gingivae. Onderstepoort Journal of Veterinary Research, 70:281–297
Bertin Thomas J. C., Thivichon-Prince Béatrice, LeBlanc Aaron R. H., Caldwell Michael W., Viriot Laurent 2018. Current Perspectives on Tooth Implantation, Attachment, and Replacement in Amniota . Frontiers in Physiology 9: 1630.
Wiersma, Kayleigh & Sander, Paul. (2016). The dentition of a well-preserved specimen of Camarasaurus sp.: implications for function, tooth replacement, soft part reconstruction, and food intake. PalZ. 10.1007/s12542-016-0332-6.
K. Wiersma, P.M. Sander. THE GRIN OF THE CHESHIRE CAT:
ISOLATED TOOTH ROWS AND OTHER DENTITION EVIDENCE FOR A RHAMPHOTHECA COMBINED WITH TEETH IN EUSAUROPOD DINOSAURS. 2019 SVP Abstract Poster.
Canudo, José,Jose, Carballido, Salgado, Leonardo,Garrido, ,Alberto. 2018/12/01, A new rebbachisaurid sauropod from the Aptian–Albian, Lower Cretaceous Rayoso Formation, Neuquén, Argentina. 63. DOI: 10.4202/app.00524.2018 Acta Palaeontologica Polonica
Hans Jakob Siber and Urs Möckli. The Stegosaurs of the Sauriermuseum Aathal. Sauriermuseum Aathal, June 2009.
Rodolfo A. García (2007) An “egg-tooth”–like structure in titanosaurian sauropod embryos, Journal of Vertebrate Paleontology, 27:1, 247-252
Kundra ́t et al., Specialized Craniofacial Anatomy of a Titanosaurian Embryo from Argentina, Current Biology (2020),
Hieronymus, Tobin. (2010). Homology and Evolution of Avian Compound Rhamphothecae. The Auk. 127. 590-604. 10.1525/auk.2010.09122.
Wang, Shuo & Stiegler, Josef & Wu, Ping & Chuong, Cheng-Ming & Hu, Dongyu & Balanoff, Amy & Zhou, Yachun & Xu, Xing. (2017). Heterochronic truncation of odontogenesis in theropod dinosaurs provides insight into the macroevolution of avian beaks. Proceedings of the National Academy of Sciences. 114. 10.1073/pnas.1708023114.
Lautenschlager, Stephan & Witmer, Lawrence & Altangerel, Perle & Rayfield, Emily. (2013). Edentulism, beaks, and biomechanical innovations in the evolution of theropod dinosaurs. Proceedings of the National Academy of Sciences of the United States of America. 110. 10.1073/pnas.1310711110.
David B Norman, Scelidosaurus harrisonii (Dinosauria: Ornithischia) from the Early Jurassic of Dorset, England: biology and phylogenetic relationships, Zoological Journal of the Linnean Society
Zaher, H., de Oliveira, L., Grazziotin, F.G. et al. Consuming viscous prey: a novel protein-secreting delivery system in neotropical snail-eating snakes. BMC Evol Biol 14, 58 (2014).























{kind=link}
{kind=link}